
Le chromosome Y : une diversité très faible intra-race bovine ?
Chez les mammifères, le chromosome Y est essentiel pour la détermination du sexe et la fertilité des mâles. Malgré son intérêt potentiel pour la sélection de mâles plus fertiles, seules quelques études se sont intéressées à la diversité génétique et à la composition génétique du chromosome Y chez les bovins. Ce manque d'intérêt pour le chromosome Y est dû au fait que les schémas d'élevage se sont longtemps concentrés sur les seuls caractères de production (lait ou viande), et ont utilisé massivement quelques taureaux à haute valeur génétique de reproduction par insémination artificielle (IA), sans autre considération.
Hormis sur une région commune très courte, les chromosomes X et Y ne recombinent pas entre eux de sorte que les haplotypes originaux du chromosome Y sont conservés au fil des générations. Leur transmission peut donc être suivie en étudiant les lignées paternelles (ou LP) dans les pedigrees. Dans cette étude, nous avons étudié la diversité du chromosome Y dans 12 races bovines et laitières élevées en France en utilisant à la fois les informations des pedigrees et des marqueurs moléculaires.
Contribution des ancêtres paternels
Lors d’une première analyse, nous avons exploité les généalogies des mâles nés entre 2015 et 2019. Pour chaque race, nous avons calculé la proportion de mâles nés d'IA par rapport à ceux issus d'accouplements naturels, ainsi que les contributions de chacun des ancêtres paternels les plus anciens enregistrés au sein de chaque généalogie, aux lignées males présentent dans la population actuelle.
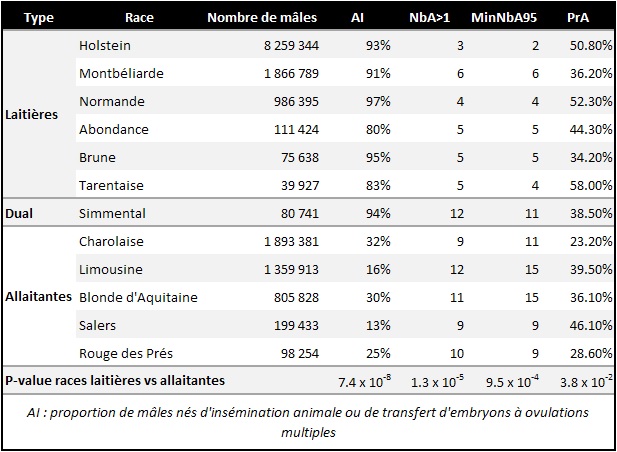
Pour étudier les lignées paternelles, nous nous sommes basés sur trois indicateurs :
- NbA > 1 : le nombre d'ancêtres mâles contribuant à plus de 1% des lignées paternelles dans la population analysée
- MinNbA95 : le nombre minimal d'ancêtres représentant 95% des lignées paternelles
- PrA : la contribution de l'ancêtre principal aux lignées paternelles
Par exemple en race Holstein, il n’y a que trois ancêtres qui ont une contribution supérieure à 1%, et parmi ces trois ancêtres, deux d’entre eux suffisent à expliquer 95% des lignées paternelles de la population des mâles nés en France entre 2015 et 2019. L’un de ces ancêtres, l’« ancêtre principal », représente 50.8% des LP de la population des mâles nés entre 2015 et 2019.
Le nombre de lignées de mâles est très faible peu importe la race. Cependant, nous avons observé des différences significatives entre les races laitières par rapport aux races mixtes et allaitantes pour tous les indicateurs étudiés. Chez les races laitières, on observe un plus faible nombre d'ancêtres contribuant à plus de 1% des lignées paternelles (NbA > 1), un plus faible nombre minimal d'ancêtres représentant 95% des lignées paternelles (MinNbA95), et une contribution de l’ancêtre principal plus élevée (PrA ; cf. tableau).
Diversité du chromosome Y
Dans un deuxième temps, nous avons évalué la diversité du chromosome Y au niveau moléculaire, en exploitant des données de séquences du projet 1000 Génomes bovins. Après avoir appliqué différents filtres, nous avons réalisé un réseau phylogénétique basé sur les informations de 811 taureaux génotypés pour 1411 SNP. Nous avons ensuite placé chaque race selon son bassin d’origine géographique (cf figure ci-dessous).

Tout d’abord, nous pouvons observer sur cette carte la présence de deux grands haplogroupes Y1 et Y2, différencié par 1266 polymorphismes et réparti selon un gradient Nord-Ouest / Sud-Est. A l’intérieur de ces deux grands haplogroupes, la plupart des branches du réseau sont races-spécifiques et reflètent la parenté géographique et génétique de notre population. Par exemple, certains taureaux de race Blonde d'Aquitaine partagent des haplotypes Y identiques avec des taureaux Limousin et Salers. On observe la même proximité entre les races Montbéliarde et Simmental ou entre Abondance et Tarentaise.
La diversité des haplotypes au sein d'une même race est plus faible que ce qui était attendu sur la base des informations généalogiques, avec des ancêtres influents indépendants portant le même haplotype du chromosome Y. Par exemple, en Normande, nous n'avons pas observé de variation entre les chromosomes Y de 30 taureaux descendant de cinq ancêtres différents. Ces résultats soutiennent l'existence d'importants effets fondateurs mâles remontant à la création des races, et donc plusieurs décennies avant l'avènement des techniques modernes de reproduction et de sélection.
En conclusion
A l'aide d'informations généalogiques et de données de séquences du génome entier, nous avons mis en évidence une diversité intra-race extrêmement faible du chromosome Y chez 12 races bovines laitières et allaitantes. Cette situation peut être expliquée par d'anciens goulots d'étranglement avant même la création des races, et serait particulièrement exacerbée dans les races laitières en raison de l'utilisation accrue des techniques de reproduction modernes. Le large ensemble de marqueurs moléculaires informatifs identifiés dans cette étude pourra permettre de futures études dédiées à l'estimation des effets des différents haplotypes du chromosome Y sur la reproduction mâle.